How Umberto Eco Explains His Thoughts on Intuition in the Text “From the Tree to the Labyrinth”
Umberto Eco, a prominent figure in the contemporary intellectual landscape as a semiotician, philosopher, and writer, addresses a wide range of themes related to the history of theories of signs and interpretation in his essay “From the Tree to the Labyrinth: Historical Studies on the Sign and Interpretation.” This collection of studies, as the title suggests, aims to explore different models through which humanity has sought to understand and organize knowledge. In this historical and theoretical context, Eco’s thoughts on intuition emerge as a crucial element for understanding his epistemology and his view of interpretive processes. Eco’s approach, deeply rooted in the philosophy of language and the study of signs, does not limit itself to an abstract examination but confronts the various concepts that intuition has assumed throughout the history of thought. The purpose of this report is to analyze in detail how Eco presents his thoughts on intuition within this important work, taking into account his broader theoretical system and the philosophical influences that shaped it.
A central element for understanding Eco’s perspective on intuition is the dichotomy between the “tree” and the “labyrinth” as representative models of our knowledge. The tree model, as illustrated by the “Arbor Porphyriana,” represents a hierarchical and finite structure of logical relationships, proceeding from the highest genus to the lowest species through a system of divisions. Eco criticizes this concept, associating it with a rigid and limited view of knowledge, similar to a dictionary that defines concepts in a closed and unambiguous way. In contrast, the labyrinth model, which is reflected in the structure of the encyclopedia, offers a more dynamic, interconnected image of knowledge characterized by potentially infinite paths. This preference for the labyrinth suggests a view of knowledge as a complex and open system, where the relationships between concepts are multiple and not rigidly predefined. This concept is close to the idea of a “rhizomatic” thought—non-systematic, fragmented, and networked, without a precise beginning and end. The transition from a “tree-like” model to a “labyrinthine” one implies a fundamental change in how we conceive access to knowledge and the role that intuition can play within these different schemes. In a hierarchical and defined system like the tree, intuition could be seen as a direct way to grasp fundamental truths or essential definitions. However, in a labyrinth of connections and interpretations, understanding more likely emerges from navigating through the network of relationships than from a simple intuitive act.
In his essay, Eco explicitly expresses a certain distrust of purely intuitive explanations, especially when it comes to understanding cultural phenomena. He emphasizes the need to seek rules and structural models to make reality intelligible, opposing interpretations based on “mystical intuitions” or a presumed “pure emanation of creative energy.” This position reflects a view of culture as a system of interactions governed by explicit laws, rather than as the product of ineffable individual intuitions. However, Eco does not completely deny the phenomenon of intuition. According to an account from one of his former students, Eco maintained that “intuition was simply fast logic,” exemplified by the deductive capacity of Sherlock Holmes. This interpretation suggests that what appears as an intuitive flash might actually be the result of a rapid and unconscious logical process based on a vast body of prior knowledge. In a more informal context, Eco himself used the expression “flash of intuition” to refer to sudden and fleeting ideas. This indicates that, although he recognizes the existence of such moments, his theoretical analysis focuses on the need to demystify intuition, trying to understand it through more structured and rational mechanisms.
Within his semiotic and epistemological framework, Eco highlights certain limitations of intuition as the sole foundation for knowledge and interpretation. In reference to the artistic field, Eco criticizes the Crocean concept of art as pure intuition, instead adhering to Pareyson’s view that describes artistic production as a process of trial and error and of patient interrogation of pre-existing matter. This perspective emphasizes that creation is not an isolated intuitive act but a process that unfolds over time through interaction with specific materials and techniques. Similarly, for Eco, creativity in general does not stem from a mysterious intuition but from the ability to recognize and activate already existing paths within the semantic organization. Eco’s insistence on the importance of codes, rules, and the “encyclopedia” as fundamental structures for understanding meaning implies that intuition alone is insufficient for navigating the complexity of semiosis. Relying solely on individual intuition would risk overlooking the intersubjective and structural dimensions that Eco considers crucial for the production and interpretation of meaning. This critique of “mystical” intuition is also linked to his caution regarding uncontrolled deconstruction, which he sees as a return to “ineffability,” similar to the inexplicability of intuition.
As an alternative or complement to intuition, Eco places concepts such as the code, the encyclopedia, and abduction at the center of his thinking. The code represents a system of shared rules that allow for the communication and interpretation of signs. The encyclopedia, a concept that Eco develops throughout his career, is understood as a multidimensional space of shared knowledge that governs the production and interpretation of signs within communicative contexts. This encyclopedia is not a simple list of information but a complex network of interconnected knowledge that constitutes a community’s cultural baggage. Finally, abduction, taken from the thought of Charles Sanders Peirce, is conceived as an inferential process through which a hypothesis is formulated to explain a surprising event. These three concepts offer a structured framework for understanding how meaning is produced and interpreted, prioritizing shared knowledge, conventional rules, and logical inference over a notion of intuition as a primary and inexplicable faculty. The emphasis on these elements suggests that for Eco, understanding and interpretation are not purely intuitive processes but activities that are based on systems of shared knowledge and rules, as well as on forms of logical reasoning.
Umberto Eco’s thought on intuition is deeply influenced by pragmatist philosophy, particularly by the works of Charles Sanders Peirce. Eco considered semiotics, which he understood as a general theory of signs, to be a fundamental part of philosophy, since the knowledge of objects and the formulation of ideas occur through signs. Eco’s approach to interpretation as a process of trial and error, similar to Peirce’s abductive reasoning, contrasts with the idea of intuition as cognition not determined by previous cognitions. Even after his reflections on cognition in “Kant and the Platypus,” which might have suggested a return to a form of pre-semiotic intuition, Eco maintained his focus on interpretation derived from hypothetical inference rather than innate intuition. Significantly, Eco expressed his disagreement with the Peircean concept of the “natural world”—the presumed innate ability to guess correctly. This divergence underscores his general caution towards purely intuitive or innate forms of knowledge, preferring an approach that emphasizes inference and cultural mediation. The influence of pragmatism, therefore, played a crucial role in shaping Eco’s view of intuition as “fast logic” and in his predilection for interpretive processes based on rules, shared knowledge, and logical inference.
In conclusion, Umberto Eco, in his essay “From the Tree to the Labyrinth,” presents a critical and reinterpretive view of intuition. While he acknowledges the subjective experience of sudden insights, he distances himself from a concept of intuition as a primary and inexplicable source of knowledge, especially in cultural and creative contexts. His preference for the “labyrinth” model over the “tree” model reflects an epistemology that values interconnectedness, complexity, and inferential processes in understanding the world. Instead of relying on “mystical” intuition, Eco emphasizes the importance of shared codes, the vast network of knowledge represented by the “encyclopedia,” and abductive reasoning as fundamental tools for the interpretation and production of meaning. His conception of intuition as “fast logic” suggests an attempt to integrate the immediacy of intuitive experience with the rational and structured processes that are at the core of his semiotic theory. The influence of Peirce’s pragmatism is evident in his emphasis on inference and the social and cultural nature of knowledge, reinforcing his reluctance to accept intuition as a fundamental and unanalyzed cognitive faculty. Ultimately, Eco’s thought in “From the Tree to the Labyrinth” invites us to reflect on the nature of knowledge and interpretation, shifting the focus from an idea of intuition as a sudden flash to a more articulated understanding of the cognitive and cultural processes that underpin our ability to make sense of the world.
Table 1: Contrast of Perspectives on Intuition
| Perspective/Thinker | View of Intuition | Eco’s Response/Interpretation |
| Croce | Art as introspective intuition | Inspired by Pareyson, art is a process of trial and error, not just intuition |
| General View | Mystical or inexplicable insight | Rejects this in favor of rules, structures, and regulated interaction |
| Eco’s Former Student | Intuition as fast logic | Seems to align with this view, exemplified by Sherlock Holmes |
| Peirce | Cognition not determined by previous cognitions | Criticizes this aspect, emphasizing inference and shared knowledge |
| Peirce | Natural world (natural instinct) | Skeptical about the idea of a mind naturally in harmony with the world |
Bibliografy
- Dall’albero al labirinto. Studi storici sul segno e l’interpretazione – Umberto Eco – Libro – IBS,https://www.ibs.it/dall-albero-al-labirinto-studi-libro-umberto-eco/e/9788893441766
- Dall’albero al labirinto: Studi storici sul segno e l’interpretazione – Google Books, https://books.google.com/books/about/Dall_albero_al_labirinto.html?id=CgUkDwAAQBAJ
- Dall’albero al labirinto. Studi storici sul segno e l’interpretazione – Umberto Eco – ebook – IBS, https://www.ibs.it/dall-albero-al-labirinto-studi-ebook-umberto-eco/e/9788893442275
- Dall’albero al labirinto. Studi storici sul segno e l’interpretazione – Umberto Eco – Libro – La nave di Teseo – I delfini | Feltrinelli,https://www.lafeltrinelli.it/dall-albero-al-labirinto-studi-libro-umberto-eco/e/9788893441766
- Dall’albero al labirinto: Studi storici sul segno e l’interpretazione eBook – Amazon.it, https://www.amazon.it/Dallalbero-labirinto-Studi-storici-linterpretazione-ebook/dp/B0716M8TTK
- Dall’albero al labirinto. Studi storici sul segno e l’interpretazione : Eco, Umberto – Amazon.it,https://www.amazon.it/Dallalbero-labirinto-Studi-storici-linterpretazione/dp/8893441764
- Dall’albero al labirinto. Studi storici sul segno e l’interpretazione – Goodreads, https://www.goodreads.com/book/show/45991248
- Dall’albero al labirinto: Studi storici sul segno e l’interpretazione – Goodreads, . https://www.goodreads.com/book/show/35163613-dall-albero-al-labirinto
- Dall’albero al labirinto: studi storici sul segno e sull’interpretazione – Goodreads, . https://www.goodreads.com/book/show/17804330-from-the-tree-to-the-labyrinth
- Umberto Eco – WE SELL ITALIAN BOOKS, . https://www.wesellitalianbooks.com/authors/umberto-eco/
- Umberto Eco, Anthony Oldcorn – Dall’albero al labirinto – Studi storici sul segno e sull’interpretazione – Harvard University Press (2014) | PDF – Scribd, . https://www.scribd.com/document/384183509/Umberto-Eco-Anthony-Oldcorn-From-the-Tree-to-the-Labyrinth-Historical-Studies-on-the-Sign-and-Interpretation-Harvard-University-Press-2014
- From the Tree to the Labyrinth by Umberto Eco | Penguin Random House Canada, . https://www.penguinrandomhouse.ca/books/792498/from-the-tree-to-the-labyrinth-by-umberto-eco/9780674049185
- (PDF) On the Problem of “Illiteracy” in the Contemporary Knowledge …, . https://www.researchgate.net/publication/287157129_On_the_Problem_of_Illiteracy_in_the_Contemporary_Knowledge_Society
- The Notion of System in the Work of Umberto Eco: Summa, Structure, Code, Encyclopaedia and Rhizome – ResearchGate, . https://www.researchgate.net/publication/351420242_The_Notion_of_System_in_the_Work_of_Umberto_Eco_Summa_Structure_Code_Encyclopaedia_and_Rhizome
- 4 Questions with Umberto … – Conversation Agent – Valeria Maltoni, . https://www.conversationagent.com/2019/09/4-domande-con-umberto-eco-autore-e-professore-emerito-universita-di-bologna.html
- We are all Umberto Eco now – Overland literary journal, . https://overland.org.au/2015/11/we-are-all-umberto-eco-now/
- Three Pragmatist Legacies in the Thought of Umberto Eco, . https://journals.openedition.org/ejpap/1232
- Eco and Pragmatism – OpenEdition Journals, . https://journals.openedition.org/ejpap/pdf/1082
- (PDF) Umberto Eco and Semiotic Creativity – ResearchGate, . https://www.researchgate.net/publication/319308005_Umberto_Eco_and_Semiotic_Creativity
- (PDF) The encyclopedia in Umberto Eco’s semiotics – ResearchGate, . https://www.researchgate.net/publication/275689765_The_encyclopedia_in_Umberto_Eco’s_semiotics
- Signs, webs, and memories: Umberto Eco as a (social) theorist – Andrea Cossu, 2017, . https://journals.sagepub.com/doi/10.1177/0725513617700414
- The Origins Of Umberto Eco’s Semio-Philosophical Project – OpenEdition Journals, . https://journals.openedition.org/estetica/7689
Kant’s Thought on the Meaning of Words
Immanuel Kant’s (1724–1804) thought is one of the fundamental pillars of modern philosophy. His critical inquiry into reason, knowledge, and morality profoundly influenced Western thought, opening new perspectives and challenging traditional conceptions. In this context, the analysis of language and the meaning of words holds a significant role, even if it is not explicitly at the center of his work. Kant did not dedicate a specific work to the philosophy of language, but his reflections on the matter are scattered throughout various writings, including the “Critique of Pure Reason,” the “Critique of Judgment,” and “Pragmatic Anthropology.”
Language and Thought in Kant
For Kant, language is strictly connected to thought and knowledge. He distinguishes between two types of language: symbolic and discursive. According to Kant, symbolic language, typical of Eastern cultures, is based on the use of images and symbols to represent reality. This type of language is considered primitive and limited, as it does not allow for the formation of abstract and universal concepts and consequently hinders cultural and moral progress. An example of symbolic language could be the use of hieroglyphics to communicate ideas and concepts.
Discursive language, on the other hand, is characterized by the use of words and the presence of a grammatical structure. This type of language, which Kant attributes to Western culture, is considered superior because it allows concepts and judgments to be expressed clearly and precisely, fostering communication and the progress of knowledge. An example of discursive language is the Italian language, with its grammatical structure and rich and articulate vocabulary.
Kant emphasizes the importance of discursive language for the development of thought and morality. Only through words is it possible to formulate universal concepts and moral principles valid for all rational beings. Furthermore, Kant argues that language, and discursive language in particular, is not innate but is acquired gradually during human development.
The Intersubjectivity of Language
A fundamental aspect of Kant’s thought on language is the emphasis on intersubjectivity. Kant argues that to formulate objective judgments about reality, thought must be intersubjectively communicable, and this is only possible through language. In other words, the ability to communicate one’s thoughts to others and to compare them with their thoughts is essential to guarantee the objectivity of knowledge.
The Meaning of Words
Intuitions and Concepts
For Kant, the meaning of a word does not lie in its simple correspondence with an external object but in its function within the system of concepts and judgments that constitute our knowledge. The word, therefore, is not a simple sign that denotes a thing but an active element that helps to structure our thought.
Kant distinguishes between intuitions and concepts. Intuitions are immediate representations of objects provided by the senses, while concepts are general and abstract representations elaborated by the intellect. For example, the sensation of the red color of an apple is an intuition, while the concept of “fruit” is a general representation that includes different types of fruits, including the apple.
Language, through words, allows intuitions to be connected to concepts, enabling the formation of judgments and knowledge of reality. The word “apple,” for example, connects the intuition of the red color, the round shape, and the sweet flavor to a general concept that allows us to classify and understand that object.
The Relationship Between Words and Concepts
In this process, the meaning of a word is determined by its relationship with other concepts and its role within the linguistic system. Kant emphasizes the importance of the correct use of language and terminological precision to avoid ambiguity and misunderstandings. For example, the word “freedom” can have different meanings depending on the context in which it is used (political freedom, freedom of thought, freedom of choice, etc.). A precise use of language requires clearly defining the meaning one intends to attribute to a word in a specific context.
A key insight in Kant’s thought is that language does not merely represent the world but actively shapes it. Words are not passive labels that we apply to objects but tools that allow us to organize and interpret reality. This view of language as an active element in the construction of knowledge distinguishes Kant from other more passive or representationalist theories of language.
Analytic and Synthetic Judgments
Another important aspect of Kant’s philosophy of language is the distinction between analytic and synthetic judgments. An analytic judgment is a judgment in which the predicate is already contained in the subject (for example, “All bachelors are unmarried men”). A synthetic judgment, on the other hand, adds new information to the subject (for example, “This apple is red”). Kant argues that synthetic a priori judgments are possible, meaning that we can know truths that are not simply deducible from the definition of concepts but that expand our knowledge of the world. This distinction between analytic and synthetic judgments had a profound influence on the subsequent development of the philosophy of language.
Analysis of Concepts and Definitions
Kant pays particular attention to the analysis of concepts and definitions. He distinguishes different types of concepts, such as empirical concepts (derived from experience) and pure concepts (derived from the a priori intellect). Furthermore, Kant analyzes different types of definitions, such as nominal definitions (which explain the meaning of a word) and real definitions (which describe the essence of a thing). This analysis of concepts and definitions helps to clarify the role of language in the formation of knowledge and to promote a precise and rigorous use of words.
The Origin of Language
Kant also questions the origin of language. He proposes an explanation that connects the origin of language to the development of human reason. According to Kant, language is not a divine gift or a product of chance but a creation of human reason that emerges from the need to communicate and share one’s thoughts with others.
The Meaning of Words in Kant’s General Philosophy
Kant’s reflection on the meaning of words fits into his general philosophy, particularly his theory of knowledge and his ethics.
Theory of Knowledge
In the “Critique of Pure Reason,” Kant argues that human knowledge is limited to the phenomenal world—that is, the world as it appears to us through the senses and the categories of the intellect. Language, as an instrument of knowledge, is also subject to this limitation. Words cannot describe the “noumenal” reality, which is reality in itself, independent of our experience. This limitation of language reflects the limitation of human knowledge itself.
Ethics
In Kantian ethics, language plays an important role in the formulation of the categorical imperative, the fundamental principle of morality. The categorical imperative, which prescribes acting according to a maxim that can be universalized, requires the use of language to express and communicate moral laws. For example, the maxim “Do not steal” can be linguistically expressed and universalized, becoming a moral law valid for all rational beings. Language, therefore, is essential for the formulation and understanding of moral principles.
Influences and Interpretations
Kant’s thought on language influenced numerous subsequent philosophers. For example, Johann Gottfried Herder, while criticizing some aspects of Kantian thought, recognized the importance of language for the formation of culture and individual identity. Ludwig Wittgenstein, in his “Tractatus Logico-Philosophicus,” also took up some Kantian themes, developing a theory of language as a “picture of the world.”
“Linguistic Kantianism” is a philosophical current of the 20th century that attributes a fundamental role to language in the structuring of thought and reality. This current, while inspired by Kant, is distinguished from his thought by its emphasis on linguistic relativism and the plurality of forms of life.
Conclusion
Kant’s thought on the meaning of words is complex and articulated. He recognizes the importance of language for the development of thought, knowledge, and morality but also emphasizes its limits and its dependence on the a priori structures of the human mind. The word, for Kant, is not a simple tool for communication but an active element that contributes to structuring our experience of the world.
Kantian reflection on language highlights a tension between the aspiration to universal concepts and the intrinsic limits of language. On the one hand, Kant emphasizes the importance of formulating universal concepts valid for all rational beings. On the other hand, he recognizes that language, being rooted in human experience, is subject to limits and conditioning. This tension reflects the difficulty of achieving universal and objective knowledge through language.
Kant’s philosophy of language had a lasting influence on subsequent philosophy, stimulating debates and interpretations that continue even today. His reflections on the role of language in knowledge, morality, and the construction of reality offer fundamental insights for understanding the nature of language and its relationship with human thought.
Charles Sanders Peirce
Charles Sanders Peirce, an American philosopher and logician, developed a complex theory of knowledge in which intuition plays a peculiar and dynamic role. Unlike Kant, who considered intuition as a passive faculty that receives information from the external world, Peirce sees it as an active process of interpretation and construction of meaning.
For Peirce, intuition is not a direct and immediate access to reality but a form of cognition that is not determined by a previous cognition of the same object. In other words, intuition is a form of knowledge that emerges from the interaction between the mind and the world and is based on the interpretation of signs and the formation of habits.
Peirce distinguishes between different types of intuition, including perceptual intuition, conceptual intuition, and abductive intuition. Perceptual intuition is the most basic form of intuition and allows us to perceive the world around us through the senses. Conceptual intuition allows us to form concepts and understand the relationships between them. Finally, abductive intuition is the most creative form of intuition and allows us to formulate hypotheses and discover new connections between things.
For Peirce, intuition is fundamental to the process of scientific inquiry, as it allows us to formulate hypotheses and interpret empirical data. However, intuition is not infallible and must always be subject to verification and revision through the scientific method.
In summary, Peirce’s thought on intuition differs from Kant’s in the following aspects:
- Nature of intuition: For Peirce, intuition is an active process of interpretation, while for Kant it is a passive faculty of reception.
- Role of intuition in knowledge: For Peirce, intuition is fundamental for the formulation of hypotheses and the interpretation of data, while for Kant it is the basis of sensory knowledge.
- Fallibility of intuition: For Peirce, intuition is not infallible and must be subject to verification, while for Kant the intuition of the a priori forms of sensibility is immediate and certain.
Furthermore, Peirce emphasizes the importance of context and interpretation in the formation of intuition. Intuition is not an isolated process but develops within a community of interpreters and through interaction with the world.
In conclusion, Peirce offers a view of intuition as a dynamic and interpretive process, based on the interaction between the mind, the world, and signs. This view differs from that of Kant, who considered intuition as a passive and immediate faculty.
Umberto Eco, as expressed in “From the Tree to the Labyrinth”
Kant considers intuition as an immediate representation of objects provided by the senses. It is the basis of our sensory knowledge and allows us to perceive the world around us. Intuition, for Kant, is passive, meaning it receives information from the external world without actively processing it. Furthermore, intuition is always spatial and temporal, as space and time are the a priori forms of sensibility—that is, the conditions that make sensory experience possible.
Eco, in his book “From the Tree to the Labyrinth,” analyzes the evolution of the concept of encyclopedia and knowledge throughout history. In this context, he criticizes the idea of intuition as a source of immediate and absolute knowledge. Eco argues that knowledge is always mediated by signs and interpretations, and that there is no direct and intuitive access to reality. For Eco, intuition is an illusion, a simplistic way of thinking that does not take into account the complexity of the cognitive process.
In summary, the following differences can be identified between Kant’s and Eco’s thought on intuition:
- Role of intuition in knowledge: For Kant, intuition is the basis of sensory knowledge, while for Eco it is an illusion that has no significant role in knowledge.
- Nature of intuition: For Kant, intuition is passive and receives information from the external world, while for Eco it is an active process of interpretation.
- Immediacy of intuition: For Kant, intuition is immediate, meaning it is not mediated by concepts or signs, while for Eco there is no intuitive and immediate access to reality.
Furthermore, Eco questions the idea of a universal and immutable order, which was the basis of the Kantian conception of intuition as the faculty of grasping the a priori forms of sensibility. Influenced by Peirce’s thought, Eco emphasizes the dynamic and interpretive nature of knowledge, which is built through a continuous process of semiosis—that is, the interpretation of signs.
In conclusion, while Kant considers intuition a fundamental faculty for sensory knowledge, Eco criticizes it as an illusion that does not take into account the complexity of the cognitive process. Eco proposes a more dynamic and interpretive view of knowledge, in line with his conception of the world as a labyrinth of signs and interpretations.
Neuroscience and Intuition
Neuroscience offers interesting insights into the neural mechanisms underlying intuition. Although intuition has traditionally been considered a mysterious and inexplicable phenomenon, neuroscientific research is beginning to shed light on the brain processes that generate it.
- The role of emotions: Intuition seems to be deeply rooted in emotions and the neural circuits that process emotional information. Brain regions such as the amygdala, insula, and ventromedial prefrontal cortex play a crucial role in this process. The amygdala processes emotional stimuli and helps to shape intuitive reactions even before we become consciously aware of them. The insula integrates bodily sensations with environmental stimuli, further contributing to intuitions. The ventromedial prefrontal cortex evaluates the meaning of emotions and helps to understand them.
- The predictive brain: The brain can be considered a “predictive machine” that constantly compares incoming sensory information with past experiences in order to predict what will happen in the future. Intuitions occur when the brain detects a significant correspondence or discrepancy between the cognitive model based on past experience and the current experience, but this information has not yet reached conscious awareness.
- Intuition and experience: Intuition can improve with experience. When a person has a lot of experience in a particular field, the brain has more information available to process the current experience and generate more reliable intuitions.
- Intuition and the right hemisphere: Some scientists believe that intuition involves different brain regions, particularly the right hemisphere, the hippocampus, and the gut. The right hemisphere is associated with creativity, emotions, and intuition.
- The role of the gut: The gut, often called the “second brain,” has an important role in emotions and intuition. The enteric nervous system, a network of neurons that lines the gut, is able to perceive and react to the environment independently of the central nervous system.
- Intuition and insight: Intuition and insight are two closely related cognitive processes. Intuition can be seen as a form of immediate and unreasoned knowledge, while insight is the ability to gain a deep and accurate understanding of a problem. Both processes seem to involve the right hemisphere of the brain.
- Intuition and flow: Intuition and flow are two cognitive phenomena that are based on the unconscious processing of information. Intuition can be considered a rapid and unconscious decision-making process, while flow facilitates this process in action, allowing one to achieve optimal cognitive control and performance without conscious deliberation.
In conclusion, neuroscience is providing an increasingly detailed picture of the neural mechanisms that underlie intuition. Intuition seems to be a complex process that involves emotions, past experience, different brain regions, and even the gut. Future research will be able to further deepen our understanding of this fascinating phenomenon.
Bibliography
- Not Those Who “all speak with pictures”: Kant on Linguistic Abilities and Human Progress – PhilArchive.https://philarchive.org/archive/LUANTQ
- Kant: Philosophy of Language – Bibliography – PhilPapers.https://philpapers.org/browse/kant-philosophy-of-language
- Kant and Herder on Reason’s Relation to Language | Kantian Review | Cambridge Core.https://www.cambridge.org/core/journals/kantian-review/article/kant-and-herder-on-reasons-relation-to-language/5BF0F1255D39BB5D11DA788643262256
- Kant’s use of the term “concept” : r/askphilosophy – Reddit.https://www.reddit.com/r/askphilosophy/comments/18i69ll/kants_use_of_the_term_concept/
- What did Kant mean by “objects” and how do they relate to intuitions and concepts?.https://philosophy.stackexchange.com/questions/82821/what-did-kant-mean-by-objects-and-how-do-they-relate-to-intuitions-and-concept
- What did Kant say on the invention of new terms? – Philosophy Stack Exchange.https://philosophy.stackexchange.com/questions/60220/what-did-kant-say-on-the-invention-of-new-terms
- Kant on Philosophy as Conceptual Analysis.https://revistas.ucm.es/index.php/KANT/article/download/91509/4564456567659/4564456694931
- Immanuel Kant – Stanford Encyclopedia of Philosophy.https://plato.stanford.edu/entries/kant/
- Technical Terms of Kantian Philosophy.https://kantphilosophy.wordpress.com/technical-terms-of-kantian-philosophy/
- Mikhail A. Smirnov, Kantian Philosophy and ‘Linguistic Kantianism’ – PhilArchive.https://philarchive.org/rec/SMIKPA-3
Researchers have used various techniques to study the neural correlates of decision-making. One common strategy is to use functional magnetic resonance imaging (fMRI) to measure changes in blood oxygen levels in the brain as people make decisions. fMRI studies have shown that the PFC is activated during decision-making, especially when the decision is difficult or complex. The PFC is also activated when people are learning from their decisions, which suggests that it plays a role in updating our knowledge about the world and how to make decisions.
Here are some of the specific brain areas that are involved in decision-making:
- The prefrontal cortex (PFC): The PFC is a large region of the brain that is involved in a wide range of cognitive functions, including attention, working memory, planning, and decision-making. The PFC is thought to play a role in representing the current state of the environment, generating possible responses, and evaluating the consequences of each response.
- Cingulate cortex: The cingulate cortex is a region of the brain that is located in the medial prefrontal cortex. The cingulate cortex is involved in monitoring the environment for threats and rewards, and it helps to prioritize decisions.
- Amygdala: The amygdala is a small, almond-shaped structure that is located in the medial temporal lobe. The amygdala is involved in processing emotions, and it helps to guide decisions based on emotional cues.
- Basal ganglia: The basal ganglia are a group of interconnected nuclei that are located deep within the brain. The basal ganglia are involved in motor control, and they help to implement decisions.
In addition to these specific brain areas, there are a number of other brain regions that are involved in decision-making, including the parietal cortex, the temporal cortex, and the hippocampus. The parietal cortex is involved in processing sensory information, and it helps to guide decisions based on the current state of the environment. The temporal cortex is involved in processing auditory and visual information, and it helps to guide decisions based on the information that is being perceived. The hippocampus is involved in memory, and it helps to store information that is relevant to decision-making.
—
The transformation of sensory input to motor output through the higher-order cognitive process of decision-making is the most challenging aspect of neuroscience.
In doing so, we are afforded a glimpse of the building blocks of higher thought and consciousness. A stimulus might motivate a particular behavior, but the action may be delayed, pending additional information, or it may never occur. This freedom from the immediacy of action means there are operations that transpire over time scales that are not immediately beholden to changes in the environment or the real-time demands of control of the body. Of course, not all decisions invoke cognition.
Many behavioral routines (swimming, walking, feeding, and grooming) have branch points that may be called decisions, but they proceed in an orderly manner without much flexibility or control of the tempo. They are governed mainly by the time steps of nervous transmission and are dedicated for the most part to particular input–output relationships.
A decision is a commitment to a proposition, action, or plan based on evidence (sensory input), prior knowledge (memory), and expected outcomes. The commitment is provisional. It does not necessitate behavior, and it can be modified. We can change our minds. The critical component is that some consideration of evidence leads to a change in the state of the organism that we liken to a provisional implementation of an action, strategy, or new mental process.
Such propositions can be represented as a plan of action: I decide to turn to the right, to leave safe shelter, to look for water, to choose a path least likely to encounter a predator, to approach a stranger, or to seek information in a book.
This view invites us to consider knowing as the result of directed (mostly non-conscious) interrogation, rather than an emergent property of neural representations.
The simplest type of decision involves the detection of a weak stimulus, such as a dim light or a faint sound, odor, or touch. Such experiments were therefore used to infer the fundamental sensitivities of a sensory system from behavior, a subfield of psychology known as psycho-physics.
The psychophysical investigation of perception began with Ernst Weber and Gustav Fechner in the 19th century. They were interested in measuring the smallest detectable difference in intensity between two sensory stimuli. It turns out they also lay the foundation for the neuroscience of decision-making because every yes/no answer is a choice based on sensory evidence.
The brain does not directly perceive a stimulus but receives a neural representation of the sample. As a result, some of the noise arises from the neural activity involved in forming this representation. It is the job of the brain to decide from which distribution the sample came, using information encoded in neural firing rates. However, the brain does not have access to the distributions, just the one sample involved in each given decision. It is the separation of these distributions—the degree that they do not overlap—that determines the discriminability of a stimulus from noise. The decision rule is to say “yes” if the evidence exceeds some criterion or threshold.
The challenge for neuroscience is to relate the terms signal, noise, and criterion to neural representations of sensory information and operations upon those representations that result in a choice.
What type of sensory information is produced by different afferent units? What is the encoding of information from the visual, auditory, and somatosensory channels?
The visual and auditory systems, in particular, employ an information-processing strategy that begins at the sensory level (retina and cochlea). The encoding allows for a significant extension of bandwidth and a transition to progressively more abstract categorization as information flows from the periphery towards the center, namely specialized cortical structures for sensory modalities: visual areas and auditory areas.
The auditory pathways: rules for encoding information
- Frequency Encoding: The auditory nerve encodes the frequency or pitch of sound. Different neurons within the auditory nerve are sensitive to different frequency ranges, allowing them to respond preferentially to specific sound frequencies.
- Intensity Encoding: The intensity or loudness of sound is encoded by the rate of firing of auditory nerve fibers. Louder sounds lead to a higher rate of firing, while softer sounds result in a lower rate of firing.
- Temporal Encoding: The timing or temporal aspects of sound are encoded by the precise timing of action potentials generated by auditory nerve fibers. Fine temporal details, such as the timing of sound onsets and offsets, are represented in the firing patterns of neurons.
- Phase Locking: The auditory nerve exhibits phase locking, which refers to the synchronization of neural activity with the phase of a sound wave. This allows the auditory system to preserve temporal information and encode the phase relationships between different frequencies.
These make perception of various auditory features, such as pitch, loudness, and timing.
The visual pathways: rules for encoding information
- Spatial Encoding: The optic nerve encodes spatial information related to visual stimuli. The retina consists of specialized cells called photoreceptors that respond to different wavelengths of light. These photoreceptors are organized in a pattern called the receptive field, which determines how visual information is spatially encoded. The activation of different photoreceptors in response to specific light patterns forms the basis for spatial encoding.
- Contrast Encoding: The optic nerve encodes the contrast between different visual elements. Contrast refers to the difference in light intensity between different parts of an image. Neurons in the optic nerve are sensitive to changes in contrast and generate action potentials based on the contrast information present in the visual stimulus.
- Temporal Encoding: The optic nerve also encodes temporal aspects of visual information. The timing of action potentials generated by neurons in the optic nerve can carry temporal information related to the onset, duration, and timing of visual stimuli. This temporal encoding contributes to our perception of motion and dynamic visual events.
- Color Encoding: The optic nerve encodes information about the colors present in a visual scene. Different types of photoreceptors in the retina are sensitive to different wavelengths of light, allowing for the discrimination of various colors. The activation patterns of these photoreceptors are transmitted through the optic nerve, providing color information to the brain.
These make perception of various visual features, such as colors, contrast, spatial position, and temporal sequence.
]]>
Le metodologie a disposizione consentono di verificare la regolare funzionalità di alcune delle principali funzioni regolatorie del cervello, anche in condizioni di anestesia generale. Si possono controllare le vie motorie, sensitive, sensoriali (acustica e visiva) e alcune risposte riflesse stimolo correlate come il riflesso “H” e le risposte “F”.
]]>
 Organization of the motor system
Organization of the motor systemWhat do we mean by “motor program” and by learning a motor skill? The central nervous system does not define in detail the displacement of the effectors (muscles), the rotations of the joints, the muscular forces, the joint pressures and the muscular activations, therefore the term engram or neuronal representation seems to provide a better indication (given the current knowledge ) of how the central nervous system codes for purposeful voluntary movements and has no relation to involuntary (automatic) movements.
Reference to “Biomechanics and motor control – defining central concepts” M.L. Latash and V.M. Zatsiorsky, 2016.
The means available to study the different functional aspects of the motor system are rather ineffective in defining a precise location of all the sub-components of the system itself. Once again the detailed study of the activity of single motor units or of the activity of single neuronal groups only provides us with specific information but does not allow us to hypothesize if not generically a motor control function. The “dynamic” approach to the control system seems to promise good prospects for understanding the motor system. In other words, only by causing a perturbation in the control system, will we be able to trace the strategies that the system is able to implement to compensate for the induced perturbation; in this way we will be able to quantify exactly how much the system is no longer able to implement due to an inefficiency linked, for example, to aging, or rather to a lesion in the system itself (in the control section or in the of the effectors).
Some questions
- What is the anatomical site of concentration of the incoming information?
- spinal cord: through the posterior horn and gamma system (gamma motor neurons)
- bulbar nuclei (gracilis and cuneate) site of the first synapse of the afferent fibers type Ia (A-alpha) from the annulospiral endings of the neuromuscular spindles, type II (A-beta) from the diffuse paranodal endings (see figure above) which make up the bundles afferent (…).
- Type II (A-beta) afferents from Pacinian bodies and paciniform corpuscles, Ib (A-alpha) fibers from Golgi tendon organs, Type III (A-delta) fibers from free endings for pain and pressuree the IV fibers (unmyelinated) from the pain-free endings and which constitute the afferent bundles (…).
- What kind of information comes from the effector periphery?
- the afferent information are proprioceptive, sensitive, pain and pressure
- What type of information transmission is encoded?
- the flow of signals through the afferent beams is characterized by a continuous discharge of frequency modulated spikes
- what are muscle synergies?
- modification of synergies over the years of life.
- which factors contribute most to the alteration of synergies
- if the experimental study with MotorBrain is particularly focused on the effect of the subject’s age as an independent factor, what parameters can be useful for identifying the alteration of motor performance (accuracy, speed and reaction time) as a function of age?
The motor system is the set of structures of the central and peripheral nervous system that allows us to transform a “nerve signal” into muscle contraction.
Works in an integrated way with the sensory and sensitive systems (visual, auditory, somatosensory and postural); the latter provide the representation of the external world, of the context in which the motor system plans, coordinates and executes the set of motor acts aimed at a purpose.
The motor system generates three types of movement:
- reflex movements: involuntary coordinated patterns of muscle contraction / relaxation in response to peripheral stimuli.
- rhythmic movements: they are repetitive and rhythmic motor activities (chewing, swallowing, scratching). They can occur spontaneously or following peripheral stimulation. The circuits that control this category of movements are located in the spinal cord and brain stem.
- voluntary movements: movements implemented with a specific purpose. They improve with practice because it allows to correct external perturbations with two strategies:
- feedback: sensory signals from the limb are used to correct the position of the limb itself
- feedforward: use of visual, auditory and tactile information to detect the perturbations that the limb will face shortly, by implementing, before the start of the movement, particular strategies based on previous experience.
Three basic laws govern voluntary movements:
- every complex movement is the result of elementary movements that have stereotyped spatial and temporal characteristics (established before starting the movement and which can be corrected by feedforward mechanisms) and are defined motor patterns.
- the reaction time depends on three elements:
- amount of information that must be analyzed in order to perform the motor task
- physical distance that the signals must travel
- stimulus modality
- The information and the motor responses correlated to them are, at least initially, elaborated in successive stages “in series”; with exercise, processing can also take place “in parallel”, increasing the efficiency of the system.
- the speed of execution of the movements is inversely related to their accuracy
Organization of the motor system
The motor system is hierarchically organized into three levels: spinal cord, brainstem, and cerebral cortex. Each level, thanks to the afferent and efferent connections, is able to organize or regulate complex motor responses. Movement information is processed by different systems, which operate in parallel.
- spinal cord: it is the lowest level of the organization. It contains monosynaptic and polysynaptic circuits that mediate reflex and rhythmic movements. Receives afferents from supraspinal centers at the level of motoneurons and interneurons that can facilitate or inhibit different populations of interneurons and coordinate movements
- brainstem: receives afferents from the cortex and subcortical nuclei; projects to the spinal cord through two systems:
- medial descending system: controls posture and integrates visual, vestibular, and somatosensitive information (vestibulospinal tract, spinal reticulum, tectospinal tract)
- lateral descending system: mainly controls the distal muscles of the limbs and the movements of the eyes and the head (rubrospinal tract)
- cortex: it is the highest level of motor organization. The areas primarily involved in movement are the primary motor area and the premotor areas.
The basal nuclei participate in the planning and execution of the movement, which together with the cerebellum are part of feedback circuits that control both the cortex and the brainstem.
The nuclei of the base are involved in motivational processes and in the choice of behavioral plans that allow adaptation to various environmental conditions.
The cerebellum is involved in the coordination and accuracy of ongoing movements and in learning motor skills.
CORTICAL MOTOR AREAS
Cortical motor areas receive cortical and subcortical inputs, process information, and emit inputs to the brainstem and spinal cord.
- Cortical afferents: from primary somatosensory area (Brodman areas 1,2,3), somatosensory association area (Brodman areas 5,7), dorsolateral frontal cortex (Brodman area 46), and contralateral hemisphere through the corpus callosum.
- Subcortical afferents: from the ventral anterior (VA) and ventral lateral (VL) nucleus of the thalamus; they carry information from the basal nuclei and the cerebellum through the formation of cortico-subcortical circuits.
The efferents are the cortico-bulbar and cortico-spinal pathways.
- the cortico-bulbar pathway controls the motor nuclei of the brain stem that innervate the facial, glosso-pharyngeal and head muscles.
- the cortico-spinal pathway controls the motor neurons that innervate the muscles of the trunk and limbs directly (synapses with the spinal motor neuron) or indirectly (synapses on interneurons with consequent simultaneous control of several muscle groups)
Anatomical and functional organization
Cortical motor areas contain neurons that project to other cortical areas and the spinal cord. The main subdivision must be made between the primary Motor areas (M1 or Brodman area 4) and the pre-motor areas (Brodman area 6).
PRIMARY MOTOR AREA
Layer V of the cortex of M1 contains upper motor neurons (Betz cells, which are very large and represent no more than 5% of the cells that project to the spinal cord) and non-Betz cells, also present in the premotor areas. The axons of these neurons descend to the brainstem through the cortical-bulbar tract and to the spinal cord through the cortical-spinal tract which, at the level of the ventral portion of the bulb form the bulbar pyramids and 90% of the fibers cross on the midline giving rise to the lateral cortical-spinal bundle; 10% of the fibers do not cross and give rise to the ventral corticospinal bundle.
The motor cortex contains a spatial map of the contralateral half of the body but not as precise as those of the somatosensory areas: a motor neuron does not correspond to a single muscle but to a group of muscles (muscle field), it follows that an M1 motor neuron codes for an organized movement. the topographical representations of movement are likely organized according to ethologically relevant categories of motor behavior. It is believed that M1 would be particularly concerned with hand and oral behaviors that occur in the “personal space”. A muscle can contract as a result of the stimulation of different cortical areas and therefore different cortical areas project (converge) on each muscle. Within the single large areas (arm, leg, etc…) there is a concentric organization: the sites that affect distal muscles are central to a larger area that also contains sites that affect proximal muscles, while sites peripheral to the central area control only proximal muscles.
The individual cortical motor neurons (Betz cells) project directly to spinal alpha motor neurons and involve more than one motor nucleus and sometimes even muscles of different joints and have phasic-tonic activity: they discharge quickly during the dynamic phase of the movement and reduce the discharge to a continuously lower (tonic) level when a stable strength level is reached.
Motor neurons of M1 code for some movement parameters:
- “lower” parameters: force that the muscles must develop (if they act against a load, they unload with a greater frequency)
- upper parameters: trajectory of the hand in the reaching movement (distinguishes them from alpha motor neurons)
There is therefore a transcortical circuit that allows M1 to elaborate rapid responses (faster than simple reaction time and less rapid than spinal reflexes) with a degree of flexibility that spinal reflexes do not have.
In the control of the movements of the fingers, the cortical neuronal populations promote mechanisms of activation and inhibition of muscles that act on all the fingers of the hand. The very nature of the motor task (power take-off or precision grip) contributes to determining the choice of cortical motor neurons to be used for a given muscle, therefore even if a spinal motor neuron is monosynaptically connected to a cortical motor cell, its activity does not it changes coherently with the variations of the cortical cell, as the multiplicity of connections that the spinal motor neuron receives makes its activity flexible, adapting to the motor task that must be performed.
Some cortical motor neurons modify their firing frequency in proportion to the force to be developed, while others reduce it. However, the latter have an excitatory function on the target muscles, but discharge only when fine movements or gradual changes in strength are performed. Their function could be a more accurate reduction of the recruitment of motor units than that which would be obtained with the simple inhibition of cortical motor neurons of the first type mentioned above.
PRE-MOTOR AREAS
They are a complex set of interconnected areas of the frontal lobe that lie anterior to the primary motor cortex and control motor functions. The cortical areas (second subdivision of Brodman) forming part of this set are: area &, area 8, area 44/45 and part of areas 23 and 24 on the medial surface of the hemisphere.
The influence on the movement is exerted through a direct route (projections to the cortico-bulbar and cortico-spinal tracts) and an indirect route through extensive reciprocal connections with M1 and therefore the efferents are similar to those of the primary motor cortex while the afferents are clearly different.
Medial and lateral pre-motor areas are distinguished:
- the medial pre-motor area deals with the initiation of movements generated by internal instructions (1 ms before the start of the movement, a negative wave appears on the EEG at the level of the medial pre-motor regions and takes the name of preparatory potential or bereitshaft potential.This area includes the SMA (supplementary motor area) involved in the process of preparing memorized movement sequences in the absence of visual cues, and the pre-SMA (presupplementary motor area) which is the main source of afferents to the SMA and projects only to it. It is involved in motor sequence learning. The pre-SMA area is activated during motor sequence learning, while the SMA is activated when previously learned movements are performed. When a motor sequence is performed efficiently and accurately, its control can pass under M1.Lesions of the medial pre-motor area in the monkey determine a reduction in the number of spontaneous movements, while they do not affect t too much on the ability to perform movements in response to external stimuli.
- the lateral premotor area deals with the selection of movements generated by external instructions. Therefore, it is involved in the stimulus-response relationship and to do so it is equipped with neurons with set-related activity (equivalent to those which in M1 encode the parameters of a movement). Its dorsal portion deals with associative learning or the execution of delayed tasks, while the ventral one adapts the attitude of the hand to the shape of the objects to be grasped and contains the “mirror neurons”. The lateral premotor area also contains, in its posterior part, Broca’s area (44 and 45 Brodman), which is essential for speech production.
Lesions of the frontal lobe in the premotor areas result in difficulty choosing a movement in response to a stimulus, although understanding of instructions and the ability to execute the movements are retained.
The pre-motor areas
They are a collection of interconnected areas of the frontal lobe that lie anterior to the primary motor cortex and control motor functions. The areas that are part of this set according to Brodman are: area 6, area 8, area 44/45 (on the lateral surface of the frontal lobe), and parts of areas 23 and 24 on the medial surface of the hemisphere.
The influence on movement is exerted through a direct route (projections to the cortico-bulbar and cortico-spinal tracts) and an indirect route (extensive reciprocal connections with M1); the efferents therefore overlap with those of the primary motor cortex, while the afferents are very different.
We can distinguish the pre-motor areas into medial and lateral.
- the medial premotor area deals with the initiation of movements generated by internal instructions (1 ms before the start of the movement, a negative wave appears on the EEG at the level of the medial premotor regions; it is called preparatory potential or ” bereit shaft potential”). This area includes the supplementary motor area (SMA), involved in the process of preparing memorized movement sequences in the absence of visual cues, and the pre-supplemental motor area (pre-SMA), which is the major source of afferents to the SMA and project only to it. Pre-SMA is involved in learning motor sequences while SMA is activated when previously learned movements are performed. When a motor sequence is performed efficiently and accurately, its control can pass under M1 (the recovery of a motor function after a lesion of M1 involves a process of learning new movements in which the SMA and probably also the pre -SMA). lesions of the monkey’s medial premotor area cause a reduction in the number of spontaneous movements, while they do not affect the ability to perform movements in response to external stimuli.
- the lateral premotor area deals with the selection of movements generated by external instructions. It is involved in the stimulus-response relationship and to do so it is equipped with neurons with set-related activity (equivalent to those which in M1 code for movement parameters). Its dorsal portion deals with associative learning and the execution of delayed tasks, while the ventral one adapts the attitude of the hand to the shape of the objects to be grasped and contains the mirror neurons (they discharge when the monkey sees grasping movements). In its rear part it also contains Brocà’s area (44 and 45 of Brodman), fundamental for the production of language. Lesions of the frontal lobe in these locations result in difficulty choosing a movement in response to a visual stimulus, although understanding of instructions and the ability to execute movements are retained.
The pre-motor areas are involved in the reaching and grasping movements: these are classic movements aimed at a goal, finalized and require a sensory-motor transformation process (processing of spatial information on the position of the object and of our arm in order to set the movement correctly). The circuits involved in reaching and grasping are parietomotor and act in parallel.
In the reaching movement, the parameters (direction and amplitude of movement) depend on the location of the target with respect to us:
- extrastriate parieto-occipital (PO) area: partly directly and partly via dorsal medial parietal (PDM) and medial intraparietal (IPM) area
- dorsal lateral premotor area (PMd)
In the grasping movement the parameters depend on the shape and size of the object:
- dorsal extra striate cortex (ES): visual information about object size
- anterior intraparietal area (IPA)
- ventral lateral pre-motor area (PMv): the type of discharge depends on the movement that the hand has to make and on the shape of the object)
Subcortical systems involved in movement
The subcortical systems involved in movement are the cerebellum and the basal nuclei.
Cerebellum
It has a homogeneous organization, but each area receives afferents from different portions of the brain or spinal cord. This suggests that the cerebellum performs the same type of computational operations on different afferent signals.
The primary function of the cerebellum is to detect the difference (“motor error”) between a planned movement and the actual movement and, once the error has been detected, send signals (through projections to the upper motor neurons), in order to induce a reduction of the same mistake.
Anatomy
It has 3 functionally distinct regions: the cerebellar cortex, the white matter, and the deep nuclei.
The cerebellar cortex is made up of three layers in which there are five types of cells. Four types of cells are inhibitory (stellate cells, basket cells, Purkinje cells, and Golgi cells). Granule cells are excitatory.
Three pairs of deep nuclei: the fastigium, the interpositon and the dentate.
The cerebellum is connected to the brain stem by three symmetrical pairs of bundles of nerve fibers, called the inferior, middle, and superior cerebellar peduncles. The latter contains most of the efferent connections of the cerebellum.
The cerebellum receives two types of afferents: mossy fibers and climbing fibers. Both are excitatory fibers (main excitatory circuit) and target Purkinje cells, but the connections they establish in the pathway are different and, therefore, generate different responses (inhibitory cortical circuits).
The cerebellum is divided into vestibulo-cerebellum, spino-cerebellum, and cerebro-cerebellum based on the origin of the afferents. The vestibulo-cerebellum relates to the lateral and medial vestibular nuclei, while the spino-cerebellum and cerebro-cerebellum relate respectively to the deep fastigium and interpositon nuclei and the dentate nucleus, through which they project to the thalamus and, finally, to the cortex .
- Muscoid fibers: has a spontaneous discharge activity that generates a constant discharge of simple spikes; if they are stimulated, the firing rate of simple spikes increases up to a few hundred spikes per second. This frequency can encode the intensity and duration of peripheral stimuli or behaviors generated by the central nervous system.
- Climbing fibres: discharge spontaneously at a very low frequency and stimuli or movement change this frequency slightly (only rarely 1-3 spikes per second are reached). These fibers possess dendro-dendritic connections which allow them to discharge synchronously; synchronous firing of climbing fibers corresponds to the appearance of complex spikes in clusters of Purkinje cells. This could be a way to signal the temporal succession of events. Climbing fibers also affect parallel (muscoid) fiber signaling; the action potentials of the former determine a moderate reduction in the effectiveness of the signals transmitted by the latter. Furthermore, the activity of the climbing fibers may result in a long-term, selective depression of the synaptic efficacy of the synchronously firing parallel fibers.
The execution of a movement determines a relative activation of climbing fibers which can generate an error signal (due to the detection of differences between expected and transmitted sensory signals), depressing the parallel fibers (which carry “incongruous” central signals) and therefore correcting the movement. As the movement is repeated the effects of parallel fiber signaling become less and less and the movement becomes more and more correct. This explains why as a result of cerebellar lesions motor learning may fail.
The vestibule-cerebellum has inhibitory efferences directed only towards the medial and lateral vestibular nuclei; in this way it controls the eye movements, the coordination of the movement of the eyes and the head and the maintenance of balance and the upright position during the walk.
Injuries of these pathways determine the inability to use vestibular information to control standing and walking: gait with an enlarged support base to compensate for the deficit that often results in falls to the ground anyway (regardless of whether their eyes are open or closed ); these patients, on the other hand, have no problems in controlling the limbs during the execution of the various types of movement when they are lying down or supported.
The spino-cerebellum emits direct efferences to the cortex, brainstem, cortico-spinal and rubro-spinal systems. Check your posture, locomotion and gaze direction. It is roughly organized in somatotopic maps, but with microscopic recordings they demonstrate a fragmented somatotopy (the afferents coming from a precise peripheral site are distributed divergently and terminate at the level of many areas in relation with circumscribed aggregates of granules). Movement control is operated with feed-forward mechanisms.
Lesions of the interpositus nucleus cause a reduction in the activity of generation of the excitatory postsynaptic potential with consequent reduced stimulation of rubro / cortico-spinal neurons, and a reduced neuronal excitability that manifests itself clinically with a reduced muscle tone (cerebellar hypotonia). The other problem is the deficit of anticipatory control on the motility of the (lower) limb ipsilateral to the lesion, the clinical manifestations of which are an oscillatory response (pendular reflexes) to an external perturbation and an irregular oscillation around the target (terminal tremor) at the end of the rejoining movement. The cerebellar dysmetria and ataxia is all the more evident the more joints are involved in the movement as the cerebellum has to mediate a greater number of proprioceptive information.
The lesions of the worm and of the nucleus of the fastygium cause alterations of the language, which becomes slow and with the emission of one word at a time (pronounced language).
The cerebro-cerebellum emits efferences towards the primary motor, pre-motor and pre-frontal cortex. It deals with the planning of complex actions, their mental repetition and the conscious evaluation of errors in movements. Injuries of this path cause decomposition of the movement of the half of the body ipsilateral to the lesion and increase in reaction times.
In general, lesions of the medial position of the cerebellum interfere with the accuracy of execution of motor responses, while lesions of the lateral portion interfere with the temporal succession of serial events.
Basal ganglia
They are the main components of the reentrant cortico-subcortical circuits that connect the cortex and the thalamus. They affect movement by regulating the activity of upper motor neurons.
Anatomy
They include four formations: the striatum (caudate, putamen, ventral striatum) and the pale globe (internal and external) emit GABAergic initorial projections, the substantia nigra (pars compacta and pars reticulata) that emits dopaminergic projections, and the subthalamic nucleus that emits projections glutamatergic excitatory.
From a functional point of view, two ways: direct and indirect.
- the direct way is through positive feedback; its activation “frees” the thalamus and cortex, facilitating movement
- the indirect path is negative feedback and its activation “inhibits” the thalamus and cortex, inhibiting movement.
The basal ganglia, relating to the cerebral cortex and the thalamus, take part in the skeleton-motor circuit. Within this circuit it is possible to identify sub-circuits organized in a somatotopic and functional way (the dorso-lateral portion of the putamen deals with the leg, the ventro-medial portion of the face and the intermediate portion of the arm). When a subject has to perform a movement and receives the indication on the direction of this movement, the cortex (M1, pre-motor, SMA) modifies the firing frequency generating the “motor attitude”, while putamen and internal pale globe modify the discharge rate to produce two possible responses:
- one is movement preparation (for neurons receiving afferents from SMA)
- the other is the execution of the movement (for neurons that receive afferents from M1)
Disturbances in the activity of the indirect route can generate hypokinetic or hyperkinetic problems. Hyperactivity of the indirect pathway causes hypokinetic disorders such as Parkinson’s disease, while hypoactivity of the same indirect pathway causes pathologies such as Huntington’s chorea.
How to design a CDSS: Clinical Decision Support System
Movement control is a process that takes place mostly autonomously (not consciously) but whose effects / results are indispensable for interaction and environmental survival. Motor behavior is the result of a long period of years of learning the motor strategies that spontaneously or specifically addressed (training), are acquired as procedural baggage.
]]>
Link al sito di Deep Learning utilities

A visualization of the model taken at one time point while running. Each sphere represents a computational unit, with lines representing the connection between units. The colors represent the output of each unit. The left and right of the image are the inputs to the model and the center is the output, which is used to guide the virtual bee down a simulated corridor. (credit: The University of Sheffield)
]]>MOTORBRAIN
Studio sull’invecchiamento fisiologico del sistema motorio
Capita frequentemente di osservare persone i cui movimenti ci sembrano impacciati, lenti o con qualche difficoltà. E il nostro giudizio è spesso influenzato dall’età della persona: più la sua età è elevata, più giudichiamo “normali” i disturbi del movimento. Questo giudizio non è però confortato da sufficienti dati quantitativi di certezza scientifica sul manifestarsi dell’invecchiamento del sistema motorio ed è quindi necessario raccogliere dati per arricchire le conoscenze sul comportamento motorio della popolazione.
La nostra ricerca si focalizza su queste problematiche e a tale scopo abbiamo sviluppato l’applicazione “MotorBrain”, che misura i movimenti di controllo della mano considerando caratteristiche quali precisione, velocità di esecuzione e tempo di reazione visivo – motorio. “MotorBrain” contiene 6 test motori che raccolgono tali informazioni. Ogni test richiede agli utenti la ripetizione di almeno tre prove, in modo da avere una migliore statistica dei dati. Inoltre, ogni test può essere eseguito con entrambe le mani. Si raccomanda di eseguire ogni test prima con la mano dominante (quella con cui si scrive o si tiene la forchetta per mangiare), poi con l’altra.
I dati raccolti dall’applicazione (questionari e movimenti degli utenti nei test) verranno inviati dall’applicazione, esclusivamente in forma anonima e in nessun modo riconducibile all’utente, ad un server universitario con una connessione sicura e salvati in un database protetto. Il database verrà usato per calcolare le distribuzioni statistiche delle misure di precisione, velocità e tempo di reazione visivo – motoria della popolazione, suddivisa per fasce di età.
Le implicazioni della ricerca sono molteplici. Dal punto di vista diagnostico, l’analisi del database potrebbe portare ad utilizzare la stessa tecnologia e metodologia dei test di prestazione motoria su pazienti con disturbo del movimento, per arrivare ad una diagnosi differenziale tra diverse patologie di “disordini del movimento” e produrre un marker precoce di “invecchiamento patologico del sistema motorio”. Dal punto di vista riabilitativo, potrebbe portare al perfezionamento di uno strumento personalizzabile ed in grado di eseguire un training riabilitativo sfruttando i processi di riadattamento funzionale del sistema nervoso centrale (plasticità), in particolare nella gestione del movimento e della memoria procedurale.
Link al sito di MotorBrain http://hcilab.uniud.it/motorbrain/
N ell’immagine a sinistra sono evidenziati i solchi e le circonvoluzioni della superficie corticale ricavate dalla RM; vi è sovrapposta l’area di attivazione sensoriale da stimolazione delle fibre del nervo mediano al polso, bilateralmente.
ell’immagine a sinistra sono evidenziati i solchi e le circonvoluzioni della superficie corticale ricavate dalla RM; vi è sovrapposta l’area di attivazione sensoriale da stimolazione delle fibre del nervo mediano al polso, bilateralmente.
Sull’emisfero di sinistra la struttura anatomica risulta deformata a causa di una lesione espansiva sottostante che, tuttavia, non impedisce una regolare attivazione afferente del sistema somato-sensoriale.
CENNI STORICI
L’idea che il cervello controlli il corpo come chi fa
muovere le marionette agendo sui fili di controllo, è suggestiva ma anche molto semplicistica. Dalla scoperta dei riflessi muscolari (riflessi osteo tendinei) nel ‘900, si è posta notevole enfasi sulla natura riflessa dei movimenti. Ivan Sechenov (1829 – 1905) e il suo studente Ivan Pavlov (1849 – 1936) hanno proposto la teoria che i movimenti, come pure le azioni mentali, siano composti da una sequenza di riflessi. William James (1842 – 1910) propose l’ipotesi della catena di riflessi, opportunamente selezionati e combinati, come motore causale dei movimenti stessi.
Gli studi sperimentali di Charles Sherrington (1857 – 1952) portarono alla teoria della locomozione come sequenza di riflessi di flessione e di estensione crociata agli arti inferiori. In seguito la teoria riflessa di Sherrinthon fu rifiutata a favore della nozione di un generatore centrale di pattern di movimenti ritmici(CPG), compresa la deambulazione.
Nel lavoro del 1935 di Nikolai Bernstein (1896 – 1966) si enfatizzò il ruolo attivo, piuttosto che solamente reattivo, dei movimenti biologici, introducendo il concetto di engramma, quale rappresentazione astratta dei movimenti a livello del sistema nervoso centrale.
Per analizzare la complessità del sistema motorio, prendiamo in considerazione due condizioni determinanti: la postura e il grasping.
La postura è caratterizzabile dalla sua configurazione e dalla sua stabilità. L’idea di un sistema di controllo basato su configurazioni di riferimento (del corpo) e di sinergie muscolari di adattamento, è l’unica al momento in grado di spiegare la complessità del movimento del corpo umano nella sua statica e nella sua dinamica.
Nel grasping le attività della mano comprendono tre sequenze :
- la selezione di uno specifico pattern di afferramento dipendente dall’obiettivo di performance e dalla forma dell’oggetto
- il movimento di raggiungimento per afferrare (reach-to-grasp)
- la manipolazione dell’oggetto esercitando forza sullo stesso, cambiando a sua posizione, orientamento e localizzazione nello spazio.
La classificazione dei movimenti di grasping comprende il power grip (l’oggetto è in contatto con il palmo della mano) e il precision grip (solo le dita sono in contatto con l’oggetto).
In letteratura sono piuttosto limitate le informazioni disponibili sulle modificazioni fisiologiche del sistema motorio indotte dal normale processo di invecchiamento. In particolare non sappiamo fino a che punto sia fisiologico, o correlato esclusivamente all’aumento dell’età, il rallentamento dei movimenti, l’incremento del tempo di reazione e la precisione dei movimenti stessi.
Recenti acquisizioni permettono di affermare la notevole importanza della capacità motoria dell’uomo anche nel processo di sviluppo / conservazione della memoria. Il cervello è un sistema di complesse interazioni con l’ambiente esterno ma anche con l’ambiente interno (il veicolo su cui è installato): si è abituato a gestire sistemi in movimento e continua a farlo fino alla fine della vita, pertanto, in attesa che i fautori del transumanesimo riescano ad installare il cervello (o meglio la nostra mente) su di un sistema semovente, noi ci limitiamo a cercare di conoscere a fondo come il cervello si adatta, nel corso degli anni, alla perdita di prestazioni del veicolo su cui è installato.
PROGETTO MOTORBRAIN
Lo sviluppo del progetto MotorBrain si è articolato proprio con l’obiettivo primario di raccogliere informazioni dalla popolazione nel più ampio spettro di età possibile, sul comportamento del sistema motorio, in particolare sulla capacità di “precision grip”.
L’app MotorBrain è stata sviluppata in collaborazione tra docenti del Laboratorio di Interazione Uomo-Macchina dell’Università di Udine e medici del Dipartimento di Neuroscienze dell’Azienda Ospedaliero-Universitaria “S.M. della Misericordia” di Udine.
ATTENZIONE: L’applicazione ed i suoi contenuti non sono in alcun modo da intendersi come sostituti di un consulto medico, diagnosi o trattamento.
In letteratura si dimostra che le capacità motorie si modificano nel corso della vita, migliorando dal bambino all’adulto e poi peggiorando progressivamente con l’avanzare dell’età [1, 2]. Non disponiamo tuttavia di misure affidabili delle performance motorie della popolazione e d’altra parte è difficile progettare uno studio di raccolta di dati “normali” in un campione esteso che possa essere rappresentativo della popolazione per estensione di età, per sesso e per prevalenza manuale.
Lo sviluppo di sistemi computerizzati di raccolta dati consente di eseguire test di misura su device collocati a domicilio o semplicemente mobili (smart phones o tablets). L’App MotorBrain consente di ottenere una rapida valutazione delle proprie performance motorie, di controllarne l’andamento nel tempo ripetendo i test nelle stesse condizioni e di fornire i dati raccolti ad un data base generale che viene costantemente aggiornato e utilizzato a scopo clini co / diagnostico nei disturbi del movimento da parte di neurologi, fisiatri e gruppi di ricerca che lavorano sulle caratteristiche del sistema motorio dell’uomo.
co / diagnostico nei disturbi del movimento da parte di neurologi, fisiatri e gruppi di ricerca che lavorano sulle caratteristiche del sistema motorio dell’uomo.
Solo di recente sono stati in parte individuati i fattori che concorrono al processo di invecchiamento fisiologico del sistema motorio [3], e il database che si sta raccogliendo permette di confrontare le performance di un paziente con sospetto disturbo del movimento e di seguirne l’andamento nel tempo in seguito ad evoluzione naturale o in seguito a provvedimenti terapeutici di tipo farmacologico o fisioterapico.
Moduli costitutivi di MotorBrain
L’applicazione sviluppata in Unity [4], è distribuita per i 3 maggiori sistemi operativi mobile (Android, iOS e Windows Phone). Sono presenti tre moduli principali che configurano due test motori ciascuno, dedicati rispettivamente alla accuratezza del movimento, alla velocità del movimento ed al tempo di reazione visivo – motorio.
Il modulo accuratezza è composto da 2 test (cerchio precisione e
quadrato precisione) in cui, utilizzando la punta del dito indice, si deve seguire la traccia colorata:
del dito indice, si deve seguire la traccia colorata:

- il primo (cerchio precisione) consiste nel seguire la banda colorata di un cerchio, con la
massima precisione possibile - il secondo (quadrato precisione) consiste nel seguire la banda colorata di un quadrato, con la massima precisione possibile
Il modulo velocità è composto da 2 test (cerchio velocità e linea zig-zag) in cui, utilizzando la punta del dito indice, si deve seguire la linea colorata nel minore tempo possibile:

- il primo (cerchio velocità) consiste nell’eseguire quanti
 più giri possibili in un tempo predefinito, rimanendo sempre s
più giri possibili in un tempo predefinito, rimanendo sempre s
ovrapposti il più possibile alla banda colorata predefinita, utilizzando esclusivamente la punta del dito indice - il secondo (linea zig – zag) consiste nel seguire il decorso della linea a zig – zag, dotata di una larghezza di banda colorata predefinita, nel minore tempo possibile, utilizzando esclusivamente la punta del dito indice
Il modulo tempo di reazione è composto da 2 test (tapping test a due posizioni e tapping test a quattro posizioni):
- il primo è un “tapping test” a due pulsanti: il compito co
 nsiste nel rispondere nel minore tempo possibile, premendo il pu
nsiste nel rispondere nel minore tempo possibile, premendo il pu
lsante che si attiva / colora, utilizzando sempre la punta del dito indice - il secondo è un “tapping test” a quattro pulsanti: il compito consiste nel rispondere nel minore tempo possibile, premendo il pulsante che si attiva / colora nelle 4 posizioni, secondo una sequenza randomizzata, utilizzando sempre la punta del dito indice
Tutti i test devono essere ripetuti in 3 prove consecutive, per consentire una validazione statistica delle misure di performance
- nel modulo accuratezza viene evidenziata l’accuratezza media delle 3 prove, espressa in unità percentuali
- nel modulo velocità viene evidenziata la velocità media delle 3 prove, espressa in cm al secondo
- nel modulo tempo di reazione viene evidenziato il tempo di reazione media delle 3 prove, espresso in millisecondi
Analisi dei risultati raccolti nel dataBase
L’obiettivo finale dell’analisi è quello di definire e costruire un modello statistico della biomeccanica del movimento di precisione (precision grip) delle dita della mano destra e sinistra. nella popolazione normale, tenendo conto dell’invecchiamento fisiologico.
Analisi preliminare dei dati raccolti
In questa fase è determinante l’esplorazione dei dati raccolti al fine di escludere le registrazioni incomplete, le registrazioni non attendibili e per eseguire uno stretto controllo dei comportamenti dei soggetti defibili come “outliers”. Tale revisione dei dati si avvale di controlli statistici di distribuzione dei dati e di controllo ispettivo dei tracciati grafici di ogni test eseguito.
Estrazione delle caratteristiche descrittive del modello
Il modello si avvale delle 3 principali caratteristiche individuate nei 3 cluster di test motori (accuratezza, velocità e tempo di reazione). Per ognuno dei 3 cluster si ottengono i seguenti descrittori di performance:
- test di accuratezza [cerchio precisione e quadrato precisione] con i seguenti parametri, media delle tre prove:
- accuratezza (1,2,3)
- tempo di esecuzione (1,2,3)
- indice di scostamento (rapporto tra percorso atteso e percorso eseguito)
- variazione temporale della velocità di esecuzione (1,2,3)
- test di velocità [cerchio velocità, cerchio precisione, quadrato precisione e la linea zig-zag] con i seguenti parametri, media delle tre prove :
- accuratezza
- tempo di esecuzione
- indice di scostamento
- velocità media di esecuzione
- numero di giri nel tempo massimo prestabilito
- test del tempo di reazione [tapping test a 2 e tapping test a 4 pulsanti] con i seguenti parametri, media delle tre prove:
- accuratezza
- tempo di reazione media
- numero di taps eseguiti
Classificazione del modello
Si utilizzano metodologie di classificazione di “machine learning” per costruire i pattern caratteristici di ciascuna classe di età dei soggetti raccolti nel database.
MotorBrain (2.0)
Nella seconda versione del progetto MotorBrain sono state introdotte particolari condizioni di attività nella gestione dei test motori che, genericamente, definiamo come perturbazioni. Tenendo conto che tali fenomeni sono inevitabilmente mediati dalla percezione visiva, si sono impostati eventi casuali di modificazione di alcune caratteristiche fisiche del test motorio, già precedentemente eseguito.
Definizione delle perturbazioni
Una perturbazione visiva nel corso della esecuzione del cerchio precisione, consiste ad esempio nella scomparsa della “traccia guida” durante l’esecuzione del compito per testare se e come il soggetto è in grado di completare il compito senza la guida sottostante.
Una perturbazione spaziale nel corso della esecuzione del tapping test, cosiste nello spostamento del bersaglio dopo un certo numero di eventi a posizione fissa.
riferimenti bibliografici
- J.S.R.Leversen,M.Haga, H.Sigmundsson, From children to adults: Motor performance across the life-span, PLoS One. 7 (2012) e38830. doi:10.1371/journal.pone.0038830
- R.D.Seidler,J.A.Bernard,T.B.Burutolu,B.W.Fling,M.T.Gordon,J.T.Gwin, et al., Motor control and aging: Links to age-related brain structural, functional, and biochemical effects, Neurosci. Biobehav. Rev. 34 (2010) 721–733. doi:10.1016/j.neubiorev.2009.10.005
- A.Cicerale, E.Ambron, A.Lingnau, R.I.Rumiati, A kinematic analysis of age-related changes in grasping to use and grasping to move common objects, Acta Psychol. (Amst). 151 (2014) 134–142. doi:10.1016/j.actpsy.2014.06.004
Nomenclatura
EMG : indica la categoria generale della metodica definita come “elettromiografia” e che attiene a tutto quanto è in relazione all’attività elettrica generata dal sistema neuromuscolare. Tale attività elettrica può essere indagata in maniera non invasiva con sistemi di registrazione di superficie (sEMG) o con metodiche invasive (elettrodi ad ago concentrico, elettrodi ad ago mono polari, elettrodi a filo) per l’analisi dettagliata delle caratteristiche delle unità motorie e delle caratteristiche di conduzione a livello della interfaccia nervo / muscolo : placca neuromuscolare.
La sEMG presenta sostanziali differenze rispetto alla EMG invasiva, in termini di valutazione quantitativa del sistema neuromuscolare. Consente la misura della forza equivalente prodotta dal muscolo, la valutazione del tempo di attivazione di un muscolo, o di un gruppo di muscoli, e la stima dell’affaticamento muscolare.
La disponibilità di sistemi di misura multicanale, consente inoltre di quantificare le sinergie muscolari in specifiche condizioni di test: coordinamento degli arti inferiori nel cammino, attività muscolare nel controllo della postura, sinergie muscolari dell’arto superiore nei movimenti di afferramento (power grip) e nei movimenti di precisione delle dita della mano (precision grip).
Sinergie muscolari
In letteratura attualmente vi sono tre definizioni di sinergia muscolare che permettono, almeno in parte, di non fare eccessiva confusione nell’utilizzo del termine sinergia muscolare:
- sinergie – A : pattern di attivazione muscolare stereotipati non pattern specifici.
- sinergie – B : pattern di attivazione muscolare che cambiano nel corso della esecuzione o in relazione alle condizioni di esecuzione.
- sinergie – C : stabilità dei movimenti specifici per una determinata prestazione / compito. L’idea di stabilità delle azioni task-specifica fu introdotta da Schoner (1995); se N variabili elementari contribuiscono a una variabile performance di M dimensioni (N > M), esiste una spazio dimensionale (N-M) in cui la variabile performance non si modifica in grandezza. Questo spazio è definito UNCONTROLLED-MANIFOLD (UCM, Scholz e Schoner, 1999). In conseguenza pure avendo una elevata variabilità inter-trial dello spazio di configurazione delle articolazioni, si ottiene una piccola variazione nel risultato e cioè nella variabile che misura la performance.
Analisi delle sinergie di tipo – C
Si tratta essenzialmente di un analisi della varianza e delle covarianze inter trial e intra trial.
Metodologie di “machine learning” nell’analisi dei segnali
In presenza di assemblaggi multicanale di segnali sEMG correlati a funzioni naturali ma complesse come le sinergie muscolari, è probabile che sia necessario utilizzare metodologie di estrazione o classificazione dell’informazione come quelle consentite dai metodi di machine learning o di approccio ai modelli classificativi con reti neurali. Si pensa ad esempio ad un approccio di “anomaly detection” che permette di individuare un pattern anomalo o patologico sulla scorta di una conoscenza sulla regolarità dello stesso fenomeno fino a quel momento osservata.
]]>La stesura di un manuale di applicazioni pratiche di metodologie strumentali destinate al controllo di procedure chirurgiche, rappresenta una sfida nella chiarezza espositiva, sulla sicurezza e sulla certezza delle informazioni che dovrebbero essere garantite all’utente, al di là di un semplicistico riferimento di tipo aneddotico. Purtroppo l’unica maniera per sviluppare il filo del discorso è quello di affidarsi all’esperienza personale, accumulata in anni di lavoro in un campo piuttosto specifico, ma che ultimamente si pone in maniera prepotente all’attenzione della equipe chirurgica e anestesiologica. Tale attenzione è motivata non solo da argomentazioni di carattere medico legale, ma soprattutto dall’enorme possibilità di approfondimento di conoscenze che la metodologia neurofisiologica intraoperatoria permette di eseguire direttamente sull’uomo.
sagittal brain
A chi è destinato il testo
Il testo è destinato alle professionalità che direttamente sono coinvolte in queste procedure e, in primo luogo, al neurofisiologo clinico che non può prescindere dall’analisi dei casi clinici presentati; in secondo luogo al tecnico di neurofisiopatologia che potrà trovare ampia documentazione delle metodiche utilizzate, delle indicazioni e dei limiti delle stesse. Non da ultime le figure del chirurgo e dell’anestesista che hanno la possibilità di confrontarsi con realtà e punti vista differenti, in particolare sugli stessi casi clinici.
Utilizzo del testo
Il manuale può essere utilizzato come testo di riferimento per la parte concettuale delle basi fisiopatologiche dei test strumentali e anche come atlante di riferimento per gli esempi e le problematiche cliniche che sono state analizzate. Lungi dall’essere completo ed esauriente, il testo vuole essere uno stimolo ad approfondire tematiche in continua e vivace evoluzione culturale. I riferimenti bibliografici sono sempre indicati allo scopo di un ulteriore approfondimento delle problematiche presentate.
Metodologie espositive
Si è cercato di conservare una metodologia espositiva quanto più omogenea tra i diversi autori, con l’ausilio di numerosi riferimenti alla propria esperienza personale.
Casistica esemplificativa
La casistica illustrata rappresenta l’esemplificazione della propria esperienza professionale, che viene esposta in quanto passibile di sicuro miglioramento, ma proprio per questo viene proposta anche come spunto di fondamentale riflessione sulle decisioni intraprese e sulle scelte effettuate in corso d’opera.
I casi descritti hanno lo scopo di passare in rassegna le diverse condizioni o complicazioni che ogni singolo caso pone; pur nella singolarità di ogni caso – paziente, si è cercato di classificare le singole problematiche per le maggiori categorie che più di frequente ci si può trovare ad affrontare.
Applicazioni della neurofisiologia intraoperatoria
E’ opinione comune che la neurofisiologia dedicata al monitoraggio sia sostanzialmente diversa da quella che si imposta e si utilizza in laboratorio. In realtà una approfondita conoscenza di tutti i principi e di tutte le metodiche che vengono utilizzate in laboratorio, è indispensabile per potere utilizzare la neurofisiologia come strumento di controllo intraoperatorio. Ad eccezione di alcune metodologie che sono applicabili solamente durante un intervento spinale o sovratentoriale in anestesia generale, tutte le metodiche utilizzate in IOM sono sostanzialmente ripetizioni o riadattamenti di quanto eseguito in laboratorio. Se ne fornisce una sintetica rassegna.
Il tronco nervoso periferico
La struttura anatomica può essere controllata ai fini di garantire un approccio chirurgico quanto più fisiologico possibile, senza danneggiare i contenuto di fibre del nervo; il controllo ideale dovrebbe estendersi alle fibre motorie, alle fibre sensitive e alle fibre del sistema termo dolorifico con le metodiche di conduzione motoria, sensitiva, risposte “F”, riflesso “H” e conduzione delle piccole fibre con stimolazione termica (laser).
Le radici spinali
Le radici spinai sono controllabili attraverso le risposte di conduzione afferente ed efferente delle fibre, a seconda del livello radicolare interessato. Si utilizzano le risposte “F”, la registrazione prossimale sui cordoni posteriori indicanti l’ingresso dell’attivazione radicolare afferente in seguito a stimolazione di uno o più tronchi nervosi derivanti dalla radice in interesse, oppure dalla superficie dermatomerica corrispondente. La stimolazione diretta della radice spinale consente di attivare risposte cmap dai segmenti muscolari innervati di competenza.
I plessi brachiale e lombosacrale
Nella chirurgia dei plessi brachiale e lombosacrale la neurofisiologia offre diverse opportunità. La stimolazione dei tronchi nervosi periferici permette di individuare le singole componenti del plesso brachiale o lombosacrale, registrando direttamente sui rami esposti del plesso. La stimolazione diretta dei fascicoli nervosi del plesso permette di individuarne l’appartenenza e la continuità anatomica, registrando le risposte cmap dai muscoli bersaglio (conduzione motoria efferente) oppure le risposte corticali da afferenza sensitiva sempre stimolando gli stessi fascicoli che contengono fibre motorie e sensitive. Anche la stimolazione elettrica diretta delle aree motorie permette di far transitare la conduzione efferente attraverso il plesso, registrando le risposte cmap dai muscoli bersaglio.
Il midollo spinale
La complessità delle vie di conduzione afferenti ed efferenti del midollo spinale può essere controllata solo in parte dalle metodiche neurofisiologiche, ma nonostante queste limitazioni, il contributo fornito non è assolutamente trascurabile. Infatti è possibile trasferire completamente le metodologie di stimolazione afferente delle vie somoestesiche dagli arti inferiori e dagli arti superiori, senza alcuna restrizione. Analogamente si ottiene facilmente l’attivazione della via motoria corticospinale e corticobulbare con stimolazione elettrica trans cranica.
La condizione operatoria (anestesia generale) ha invece consentito di sviluppare la metodica di registrazione della onda “D”, attraverso l’applicazione nello spazio sub durale o epidurale, di un catetere spinale registrante in sede prossimale e distale rispetto alla sede della lesione. In tal caso si ha il vantaggio di non dovere registrare dal muscolo bersaglio e pertanto il paziente può anche essere curarizzato e si evitano in tal modo gli artefatti da movimento muscolare che possono interferire con la stabilità del campo operatorio sotto l’ingrandimento microscopico.
Il tronco cerebrale
Numerose strutture fisiologicamente complesse racchiuse in un piccolo spazio anatomico, sono altamente suscettibili agli insulti di tipo meccanico (compressione / infiltrazione) e di tipo vascolare. La neurofisiologia ci consente di controllare la conduzione delle vie afferenti somoestesiche del lemnisco mediale, le vie afferenti uditive (lemnisco laterale), le vie efferenti attraverso il fascio corticospinale e corticobulbare e alcune risposte riflesse come il riflesso di ammiccamento che coinvolge le branche afferenti del V nervo cranico e le vie efferenti del VII nervo cranico. Le risposte riflesse prendono origine da una serie di connessioni multi sinaptiche tra i nuclei del VII e del V nervo cranico, omo e contro laterali. Tutte queste possibilità di esplorazione possono essere direttamente trasferite dal laboratorio alla sala operatoria, senza alcuna modifica, se non in senso positivo di rendere l’informazione acquisita bilaterale e di utilizzare una stimolazione trans cranica elettrica anziché magnetica, in quanto il paziente è in anestesia generale.
nervo cranico muscolo target
III – oculomotore retto mediale dell’occhio
IV – trocleare obliquo superiore
VI – abducente retto laterale dell’occhio
V – trigemino massetere
VII – faciale orbicolare dell’occhio, orbicolare della bocca, temporale
IX – glossofaringeo miloioideo, palato molle
X – vago corde vocali
XI – accessorio spinale trapezio, sternocleidomastoideo
XII – ipoglosso emilingua, palato molle
In aggiunta a quanto eseguibile anche in laboratorio, si dispone della possibilità di documentare la conduzione efferente dei nuclei dei nervi cranici rispettivamente collocati a livello del bulbo, del ponte e del mesencefalo. Nella tabella sono riassunti i nervi cranici ed i rispettivi muscoli bersaglio da cui è possibile documentare una risposta di potenziale d’azione composto prodotta dalla stimolazione delle fibre del nervo oppure dalla struttura cellulare componente il nucleo motore del nervo. Ponendo degli elettrodi registranti sul muscolo e stimolando il nervo corrispondente si ottengono risposte cmap che da un lato confermano la specifica competenza di innervazione del nervo stimolato e dall’altro permettono di individuare, nel caso si disponga di un sistema di registrazione multicanale, lo specifico nervo nel caso di più strutture ravvicinate come a livello pontino.
La fossa cranica posteriore
Negli interventi in fossa cranica posteriore abbiamo a disposizione tutto l’insieme di metodologie descritte nel caso del tronco cerebrale, cui dobbiamo aggiungere alcune considerazioni. Ad esempio non viene nemmeno presa in considerazione la possibilità di controllare alcune funzioni del cervelletto coinvolto da lesioni emisferiche o della porzione vermiana. Ma altrettanto importante sarebbe il controllo delle funzioni emisferiche cerebellari quando si verificano compressioni chirurgiche come l’applicazione di spatola di appoggio per aprire una strada a livello dell’angolo ponto cerebellare. Altrettanto importante sarebbe poter conoscere l’eventuale rischio di insorgenza di atassie assiali o lateralizzate in seguito a danno ischemico cerebellare.
Le strutture sovratentoriali
Negli interventi sulle strutture sovratentoriali il compito del monitoraggio intraoperatorio si arricchisce di un’ampia area di competenze, in particolare quando l’intervento viene condotto in anestesia locale a paziente sveglio e collaborante (awake craniotomy).
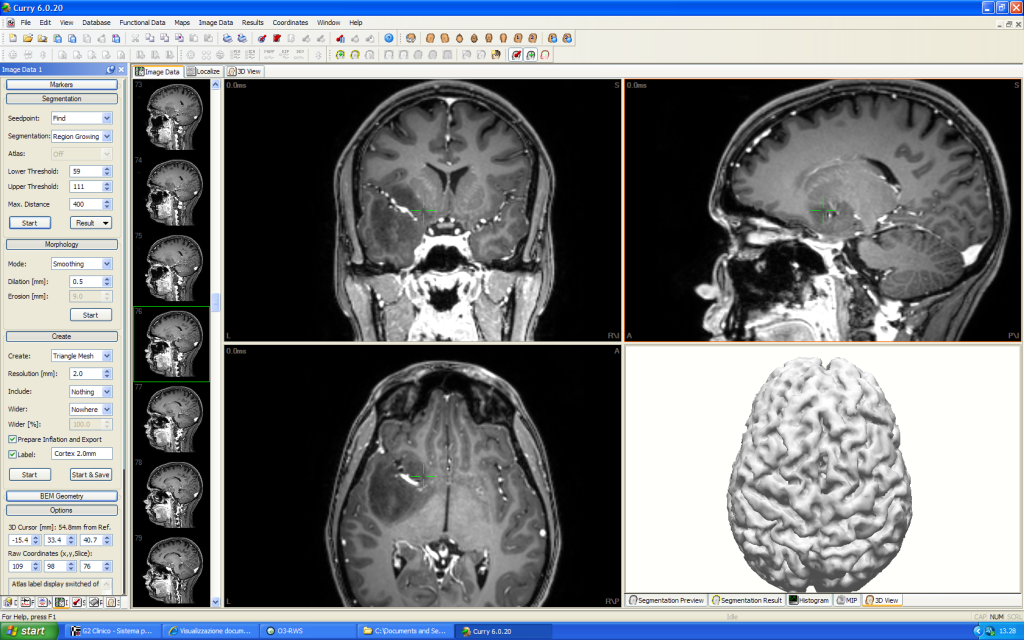
Negli interventi in anestesia generale è giocoforza che il controllo neurofisiologico sia centrato sul controllo delle vie di conduzione afferenti ed efferenti. La nostra esperienza tuttavia ci permette di raccomandare anche un controllo dell’attività elettrica spontanea (Eeg) sia con elettrodi posti sullo scalpo, si con elettrodi disposti sulla superficie corticale esposta e in sede sub durale. Si sottolinea che spesso è importante il controllo non solo della via cortico spinale e cortico bulbare e della via afferente somoestesica, ma anche della afferenza visiva, in particolare quando le lesioni temporali si portano molto in profondità fino a raggiungere le radiazioni ottiche.
In particolare il monitoraggio intraoperatorio del sistema motorio, in questi ultimi anni ha ricevuto notevole impulso e innovazione con l’introduzione della possibilità di registrare direttamente il segnale di conduzione della via motoria spinale (cordoni anterolaterali). Tale metodica è piuttosto insensibile alle variazioni dell’anestesia generale ed linearmente correlata all’intensità di stimolazione applicata a livello corticale (Amassian VE, 2002), al contrario della risposta muscolare cmap che presenta fluttuazioni caratteristiche dell’attivazione di una seconda stazione nucleare che è il motoneurone spinale (ciò comporta una relazione non lineare con lo stimolo elettrico applicato al motoneurone corticale).
L’onda “D”, o diretta , ottenuta da una singola stimolazione elettrica corticale, è il segnale di conduzione delle fibre del tratto cortico spinale, registrabile sulla superficie piale del midollo spinale. Tale componente è condotto dalle fibre cortico spinali a rapida velocità di conduzione senza interposte sinapsi. In relazione alla tipologia e intensità di stimolazione, in coda all’onda “D” si possono aggiungere componenti indirette (cosiddette onde “I”) che sono il risultato di un’attivazione transinaprica di interneuroni che scaricano lungo la via cortico spinale. La forma d’onda del segnale è tipicamente trifasica (pos – neg – pos) se biene registrata in modalità monopolare (elettrodo attivo in prossimità delle fibre ed elettrodo di riferimento distante) oppure bifasica (neg – pos) se registrata in modalità bipolare (coppia di elettrodi entrambi applicati sulla superficie spinale delle fibre attive).
La morfologia dell’onda “D” esprime l’entità del sincronismo di conduzione delle fibre (contengono differenti velocità di conduzione e, pertanto, all’aumentare della distanza di conduzione si accentua l’entità della de sincronizzazione delle stesse; e viceversa, a parità di distanza di conduzione, la comparsa di una de sincronizzazione di segnale è indicativa di insorgenza di blocco di conduzione fisiologica o patologica di sottopopolazioni di fibre ); fornisce un’indicazione quantitativa del numero di fibre attive. (ulteriore approfondimento nel capitolo spinale)
La trasformazione del segnale dell’onda “D” da trifasica a monofasica è indicativa della comparsa di una lesione parziale delle fibre del fascio cortico spinale. Questo fatto consente di individuare la sede della lesione prodotta da una contusione traumatica o da una compressione tumorale delle fibre del midollo spinale utilizzando multipli elettrodi di registrazione subdurali o epidurali.